
 服务热线:0571-83782130服务热线:0571-83782130
服务热线:0571-83782130服务热线:0571-83782130
NEWS
新闻中心
SERVERS
服务项目
NEWS
最新资讯
发布时间:2021-06-11
核心摘要
肢端肥大症是一种由生长激素(GH)分泌过多引起的病理疾病,主要由垂体腺瘤产生。长时间暴露过量的生长激素会导致广泛的系统表现和共病,研究过量生长激素对细胞水平的影响可以帮助了解肢端肥大症健康并发症和共病的潜在原因。在之前的研究中,已经证明,在肢端肥大斑马鱼模型中,过量的GH减少了体侧种群(SP)干细胞,并诱导了过早衰老的迹象。
在本次研究中,我们在基因表达水平上更深入地研究肢端肥大症的衰老及肢端肥大症是否在不同器官诱导衰老遗传特征。在肢端肥大症和老年斑马鱼的不同器官中,我们发现了重叠的差异表达基因(DEG)。大约一半的肌肉、肝脏和大脑的肢端肥大症基因与老年斑马鱼的DEG重叠,我们确定了重叠DEG的生物学功能。富集数据库分析和基因本体研究表明,大多数重叠的肌肉基因参与衰老代谢,而重叠的肝脏基因参与代谢途径、缺氧反应和内质网应激。因此,本研究在基因表达水平上提供了肢端肥大症的完全老化遗传特征。

一、引言
丙激素肥大是一种由过量生长激素(GH)水平以及随后的胰岛素样生长因子1(IGF-1)引起。乙囊肿大通常由垂体腺瘤引起。在较少的病例中,该疾病可能是由异位生长激素分泌或垂体增生引起的。巨人症和肢端肥大症都是由过量的GH和IGF-1分泌物引起的罕见疾病;然而,巨人症发生在青春期前生长激素过量触发线性发育时,而肢端肥大症发生在附生印章关闭后。
过度暴露于生长激素会导致身体毁容,包括宽阔的双臂、腿部、增厚的软组织、脸变大、鼻子和前额突出。渐进性肢端肥大症的特征是骨骼缺陷导致背后凸和胸腔变形。
除了异常的软组织生长和肥大外,肢端肥大症患者还出现了许多系统性表现和共病,包括胃肠道疾病、生殖疾病、关节炎、腕管综合征、虚弱无力、结肠息肉、糖尿病、肾病、神经病变和心血管疾病。
在过去的二十年里,我们对巨人症和肢端肥大症的遗传病因学有了更好地了解。几种新的基因突变会破坏它们细胞内介导生长激素素分泌过多的生长营养途径,包括芳烃受体相互作用蛋白(AIP)基因种系突变和X连锁生长激素综合征(XLAG)性状。
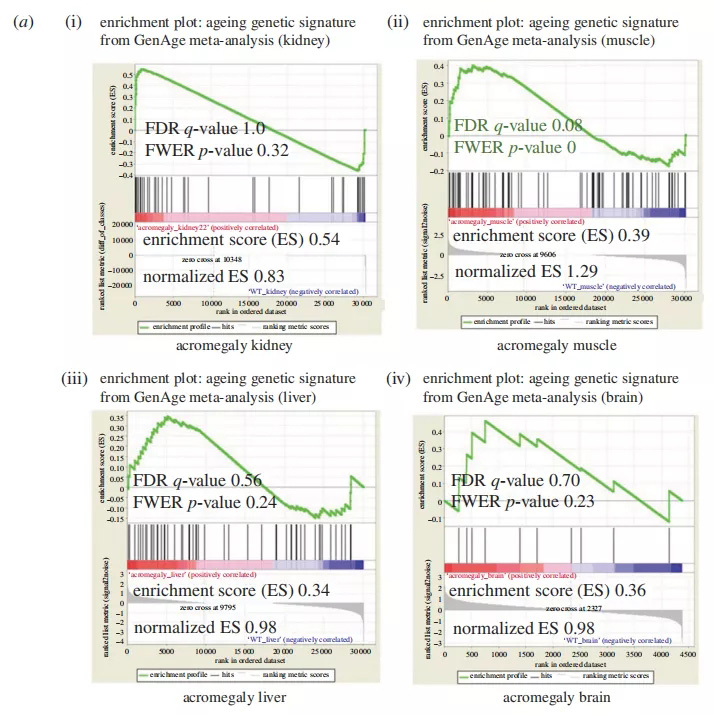
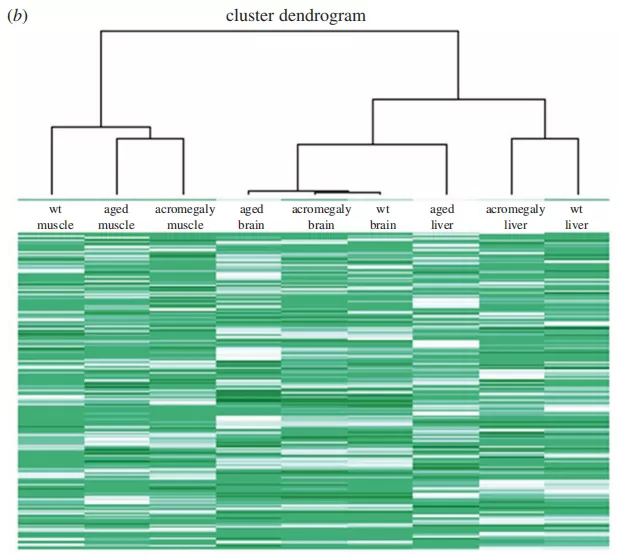
图1所示,肌肥大肌肉在基因表达水平上老化。(a)肢端肥大症肾、肌肉、肝脏和大脑中基因特征基因老化的GSEA结果。小于0.05的显著p值和小于0.25的FDRq值以红色表示。(b)层次簇分析来自WT肌肉、肝脏和大脑、肢端肥大症模型(1岁)和老年斑马鱼(3岁)的树状图和RNA序列数据热图。
肢端肥大症的全身表现广泛遍布全身,表明过量生长激素的病理作用作用于细胞水平。发现肢端肥大症的潜在遗传原因是至关重要的开发新的诊断和治疗方法。然而,较少的研究集中在过量生长激素的病理影响,以及生长激素如何在细胞水平上起作用,导致肢端肥大症合并症。
在之前的研究中,我们开发了一个过度分泌的肢角肥大斑马鱼模型,以研究过量GH对细胞生物学的病理影响。在肢端肥大症的体细胞和干细胞中,已经发现了许多被破坏的参数,如DNA损伤的增加和DNA修复途径的受损。在我们的肢端肥大症模型中,由于生长激素过多而引起的主要疾病包括体内侧种群(SP)干细胞数量的逐渐减少和干细胞氧化应激的增加。此外,我们还观察到了过早衰老的迹象。
因此,在这里,我们通过研究肢端肥大症是否在不同器官中表现出丰富的衰老基因特征来研究在基因表达水平上更深入的衰老。此外,我们研究了肢端肥大症和衰老差异表达基因(DEGs)之间是否存在重叠,并确定了肢端肥大症和衰老之间共同的生物学主题。
二、结果
2.1 基因表达水平老化
在我们之前的研究中,罗非鱼过表达斑马鱼肢端肥大模型,以研究过量生长激素对细胞生物学的病理影响。我们已经在细胞确定了细胞水平上许多被破坏的信号途径和生物过程。主要的发现是在我们的模型中干细胞数量的下降和衰老的过早迹象。
为了研究肢端肥大症在基因表达水平上的衰老,我们使用了基因年龄数据库,以确定不同器官衰老的共同遗传特征。我们使用了之前发表的从野生型肝脏、肌肉、肾脏和大脑(WT)(1岁)、肢端肥大症(1岁)和老年斑马鱼(3岁)提取的mRNARNA序列数据(基因表达综合(GEO)加入号RNAsEq数据。gse113169)。使用来自肝、肌肉、肾脏和大脑的肢端肥大症RNAseq数据进行基因集富集分析(GSEA),以检测相应器官中基因基因特征基因的富集。
如图1a中可见,所有肢端肥大器官的老化遗传数据集的富集分数呈正,表明各器官基因基因集的上调;然而,富集仅在肢端肥大肌肌肉中显著(p<0.01和错误发现率(FDR)q值=0.08)。
为了证实这些结果,我们用不同的方法研究了基因表达水平上的肢端肥大症衰老。我们从野生型WT品系的肌肉、肝脏和大脑、肢端肥大模型(1岁)和老年斑马鱼(3岁)生成了RNAseq数据的分层树状图(图1b)。重要的是,层次聚类产生了相同的结果,并在肢端肥大症和老年斑马鱼的基因表达模式上仅在肌肉中,但在肝脏或大脑中没有相似性(图1b)。


图2. 肢端肥大症和老年肌肉基因与WT肌肉图。Y轴,−log10(p值);X轴,log2(折叠变化)。橙色的基因被认为是显著的(n=3)。
2.2 肢端肥大症和老年斑马鱼基因表达差异
p值之间的过度表达基因用于识别不同器官中肢端肥大(1岁)和年龄斑马鱼(3岁)的DEG(=Adj p<0.05)。
图2显示了肢端肥大症和老年肌肉、肝脏和大脑的RNAseq基因的火山图。
由于我们的肢端肥大症模型显示出过早衰老的迹象和与老年肌肉相似的基因表达模式,我们研究了肢端肥大症的DEGs是否与老年斑马鱼的DEGs重叠。我们建立了肢端肥大症和老年肌肉、肝脏和大脑的上下凹陷的维恩图。

图3. 乙度重叠 (a)肢端肥大症和老年斑马鱼肌肉、大脑和肝脏上下图的静脉图。(b)柱形图,显示上下肢端肥大症密度与老年斑马鱼重叠的百分比
虽然老年斑马鱼的DEG数量较高,但大多数肢端肥大症DEG与老年DEG重叠(图2和3a)。值得注意的是,也以同样的方式观察到重叠;肢端肥大症向上的凹陷与老化的凹陷重叠,而不是向下的凹陷,反之亦然(图3a)。
超过50%的肢端肥大症下降指数与肌肉中的老年指数重叠,而约40%的上升指数与老年指数重叠(图3b)。重要的是,肢端肥大下降和老年下降之间或肢端肥大上升和肌肉老年下降之间几乎没有重叠(图3a)。这些结果支持了我们之前的观察结果,即肢端肥大肌在基因表达模式上与老年斑马鱼相似。
虽然层次聚类和GSEA结果(图1)并没有显示肢端肥大症大脑的老化,但Venn图显示,65%的肢端肥大症下降的DEGs与大脑中老化的DEGs重叠。约40%以上密度在老年中重叠(图3)。有趣的是,在肝脏中也观察到同样的结果(图3)。
2.3 丰富了生物主题和肌肉中重叠凹陷的信号途径
因此,我们对重叠的DEGs进行了富集分析,以确定肢端肥大症和衰老之间的共同信号途径和生物过程。肢端肥大症与老年肌肉之间重叠DEG的基因本体(GO)分析显示,生物过程中代谢途径如乙醇、氧酸和类固醇代谢的富集(图4a),以及分子功能酶活性的富集(图4a)。我们使用KEGG数据库对丰富酶活性的作用进行了分类。分析表明,大部分富集酶活性与代谢过程有关,而细胞成分分析显示球蛋白-血红蛋白复合物的富集(图4a)。
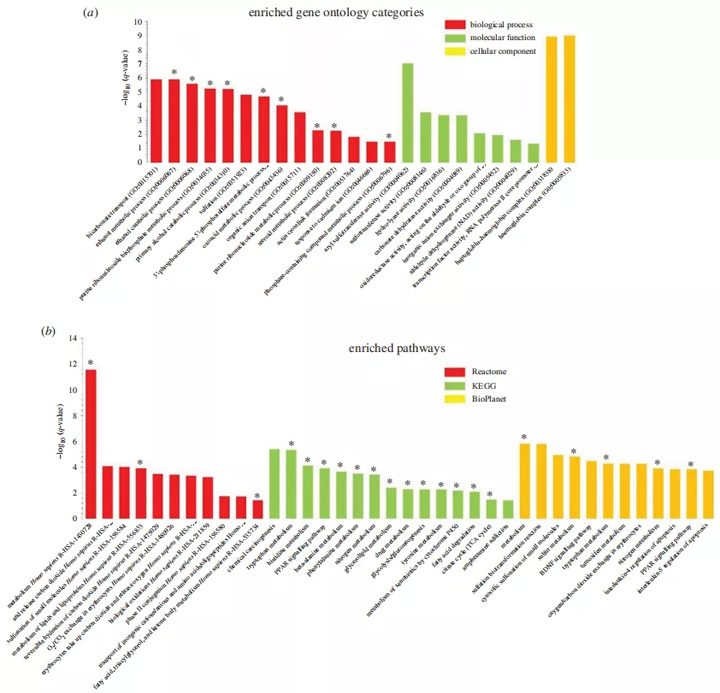
图4 丰富的肌肉重叠障碍的途径分析。(a)围棋分析显示了丰富的生物过程、分子功能和细胞成分。(b)来自反应组、KEGG和生物行星数据库中的肌肉重叠DEGs的丰富信号通路的直方图。y轴,富集的统计学意义;x轴,途径类别。与代谢相关的途径和GO类别用星号表示。BDNF,脑源性神经营养因子;PPAR,过氧化物酶体增殖体激活受体。
有趣的是,通路分析表明,大多数富集的信号通路也与代谢有关(图4b)。反应组数据库分析揭示了脂质、磷脂、脂肪酸和甘油三酯的代谢途径的富集,而KEGG和生物行星数据库分析显示脂肪酸、糖酵解、氮和各种氨基酸代谢途径。
为了研究参与肌肉老化代谢的重叠基因的百分比,我们将代谢相关基因量化为重叠基因的总数。我们发现,大多数与代谢相关的重叠DEGs都被下调了。大约50%的向下重叠DEG和20%的向上重叠DEG与代谢相关(图5)。

图5 显示与肌肉代谢过程相关的上下重叠密度的百分比的饼图
2.4 丰富的肝脏重叠病的生物主题和信号途径
维恩图显示,约40%的肢端肥大症肝脏DEGs与老年DEGs重叠(图3)。因此,我们研究了这些重叠基因的生物学功能。
对肝脏中重叠DEG的分析显示,三类主要富集:代谢途径、缺氧反应和E室应激。对缺氧的反应、脂肪酸代谢过程、转录调节、蛋白激酶R(PKR)样内质网激酶(PERK)介导的未展开蛋白反应和ER都是丰富的生物主题(图6a)。

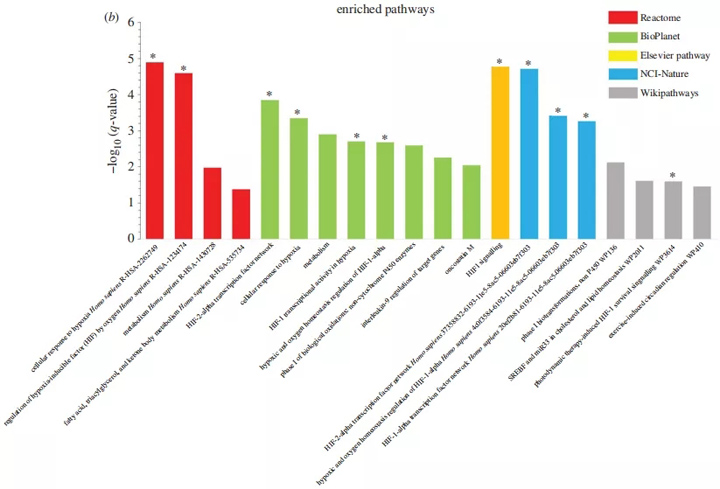
图6 富集转移和肝脏重叠病的途径分析。(a)GO分析显示肝脏重叠DEG的丰富的生物过程、分子功能和细胞成分。(b)来自反应组、爱思唯尔通路、维基通路、NCI自然和生物行星数据库的肝脏重叠病的丰富信号通路的直方图。y轴,富集的统计学意义;x轴,途径类别。与缺氧相关的途径和GO类别用星号表示。HIF-1,缺氧诱导因子;miR33,微RNA标识符;SREBF,甾醇调节元件结合蛋白基因
同样,分析显示,与缺氧和代谢相关的途径丰富(图6b)。重要的是,GSEA对肢端肥大症(1岁)和老年斑马鱼(3岁)的肝脏缺氧的反应显著下降(图7a)。
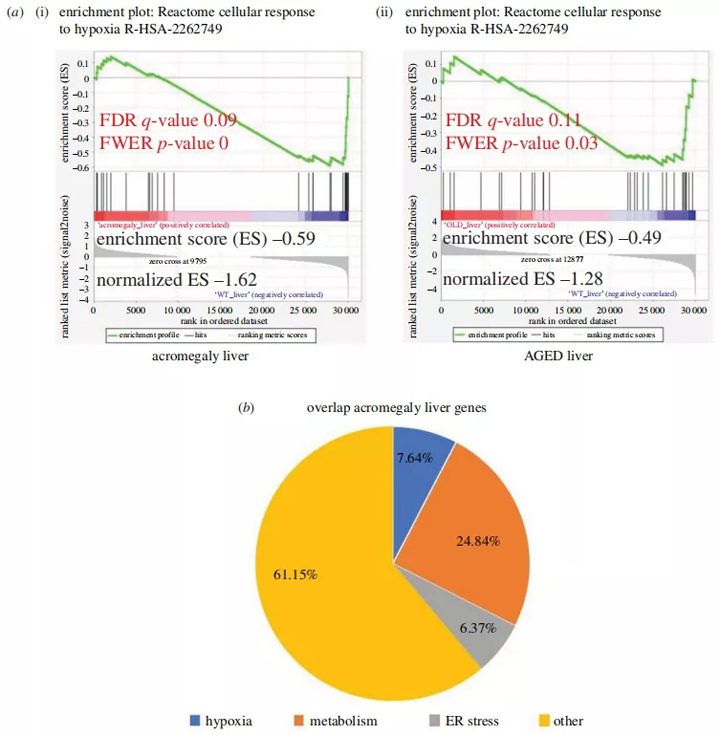
图7 重叠的肝脏DEGs中的缺氧、代谢和应激相关基因。(a)细胞对缺氧反应的统计学上显著的GSEA结果的说明。显著的p<0.05和FDRq值<0.25都是用红色书写的。报告的p值为0.0,表示实际的p值小于0.01(n=3)。(b)显示与肝脏代谢、缺氧和急诊室应激相关的重叠密度百分比的饼图。
因此,我们将与代谢、缺氧反应和急诊室应激相关的重叠基因量化为肝脏中重叠基因的总数。我们发现肝脏中大多数重叠基因与这三类相关(图7b)。
2.5 丰富的生物主题和大脑中重叠障碍的信号途径
与肌肉和肝脏相似,维恩图显示,大多数肢端肥大症脑DEGs与重叠老化的大脑。如图3所示,大约65%的肢端肥大症下降与大脑中的下降障碍重叠。
对大脑中重叠DEG的分析表明,转录、DNA结合、神经营养蛋白结合和神经递质转运的积极调节丰富了生物学过程和分子功能。细胞成分富集肌钙蛋白、肌钙蛋白和皱膜的细胞突出(图8a)。
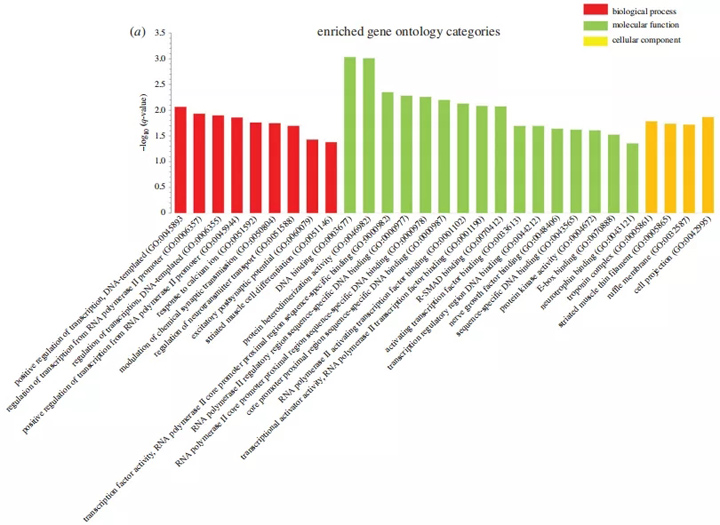

图8 丰富的围棋和大脑重叠障碍的途径分析 (a)围棋分析显示丰富的生物过程、分子功能和大脑重叠的细胞成分;(b)来自反应组、KEGG和生物行星数据库的大脑重叠DEGs的丰富信号通路的直方图。y轴,富集的统计学意义;x轴,途径类别。cGMP-PKG、环鸟苷一磷酸依赖蛋白激酶或蛋白激酶G、epo、促红细胞生成素、ERKs、细胞外信号调节激酶、FOS、Fos原癌基因、GM-CSF、粒端集落刺激因子、HTR1-5-羟色胺受体1、JNK、c-Junn-n-n端激酶、有丝分裂原活化蛋白激酶、NGF、神经生长因子、NMDA、n-甲基-d-天冬氨酸、R-SMAD、受体调节SMAD、TRK、原肌球蛋白受体激酶。
反应组、KEGG和生物行星数据库的路径分析显示,丰富了发育生物学、间隙结、昼夜节律夹带、长期增强和MAPK信号途径(图8b)。
2.6 生长激素培养的人类卵母细胞在基因表达水平上显示出衰老
在肢端肥大斑马鱼模型中,过量的生长激素影响了衰老、干细胞数量和完整性。因此,研究过量的GH是否会对人类细胞有同样的影响是很重要的。在这里,我们已经尝试着学习GEO(基因表达的综合)。
我们使用GSEA分析来研究转基因卵母细胞是否丰富了衰老特征。有趣的是,这个基因集和一个针对过早衰老疾病的数据集(沃纳综合征,MSigDBno。在GH培养的人卵母细胞中显著升高(图9)。
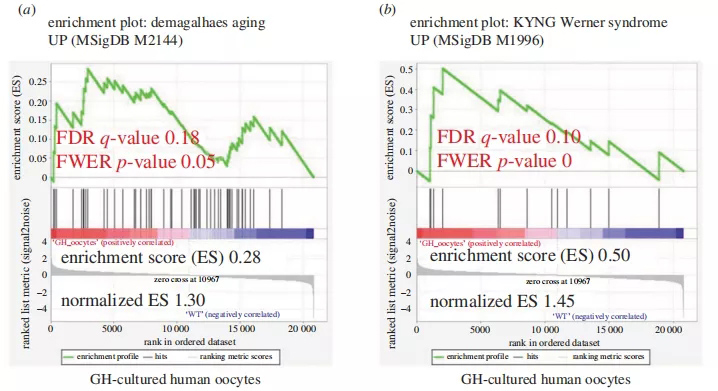
图9 GH培养的人卵母细胞在基因表达水平上显示出老化 (a)基于对微阵列数据的元分析,对随年龄持续过表达的基因组集进行统计学上显著的GSEA结果的说明;(b)过早衰老障碍数据集(Werner综合征,MSigDB号。M1996)。显著的p<0.05和FDRq值<0.25都是用红色书写的。报告的p值为0.0,表示实际的p值小于0.01。
我们使用GSEA分析来研究转基因卵母细胞是否丰富了衰老特征。有趣的是,这个基因集和一个针对过早衰老疾病的数据集(沃纳综合征,MSigDBno。在GH培养的人卵母细胞中显著升高(图9)。
三、讨论
斑马鱼是研究人类遗传疾病的一种很有前景的模式生物,因为它有完整测序的基因组、易于遗传操作、幼虫透明、生命周期短和成本低。此外,斑马鱼在生理学上与哺乳动物相似。
为了确定肢端肥大症合并症背后的分子机制,一种罗非鱼-GH过表达的斑黄鱼被开发为肢端肥大症模型。我们已经证明,过量的GH会影响到全身的细胞完整性。
此外,我们在肢端肥大症体细胞和干细胞中发现了许多破坏的生物参数。有趣的是,我们的斑马鱼模型显示了肾脏、肌肉和大脑中过早衰老和SP干细胞减少的迹象。这鼓励了我们在基因表达水平上更深入地研究肢端肥大症的衰老。
参考资料:
[1] Petrossians P et al. 2017 Acromegaly at diagnosis in 3173 patients from the Liège Acromegaly Survey (LAS) database. Endocrine-Related Cancer 24, 505–518.
[2] Hannah-Shmouni F, Trivellin G, Stratakis CA. 2016 Genetics of gigantism and acromegaly. Growth Hor. IGF Res. 30, 37–41.
[3] Elbialy A, Asakawa S, Watabe S, Kinoshita S. 2018 A zebrafish acromegaly model elevates DNA damage and impairs DNA repair pathways. Biology 7, 47.
[4] Colao A, Grasso LFS, Giustina A, Melmed S, Chanson P, Pereira AM, Pivonello R. 2019 Acromegaly. Nat. Rev. Dis. Primers 5, 20.
文丨Abdalla Elbialy
翻译丨梁晓晴
DOI丨Elbialy A. 2021 Ageinggenetic signature of hypersomatotropism. OpenBiol. 11: 200265.