
 服务热线:0571-83782130服务热线:0571-83782130
服务热线:0571-83782130服务热线:0571-83782130
NEWS
新闻中心
SERVERS
服务项目
发布时间:2021-05-14
文丨Kristin Dietrich, Imke AK Fiedler,Anastasia Kurzyukova,Alejandra C López-Delgado,Lucy M McGowan, Karina Geurtzen,Chrissy L Hammond,Björn Busse,and Franziska Knopf
翻译丨郭胜亚
发表于《Journal of Bone and Mineral Research》, Vol. 36, No. 3, March 2021, pp 436–458.

核心摘要
斑马鱼是硬骨鱼(bony fish),与哺乳动物有共同的祖先,属于骨鱼门,它们的骨骼系统继承自骨鱼门。事实上,硬骨动物和哺乳动物在骨骼元素、骨化机制和骨基质成分方面有许多共同的遗传保守特征。
然而,在骨骼研究中对斑马鱼进行研究时,需要考虑与骨形态和功能相关的差异。本文就斑马鱼的骨骼结构、脊柱形态及相关解剖结构进行综述。我们概述了斑马鱼不同的骨化类型和骨细胞,并在显微组织水平描述骨基质组成,重点评估矿化。正如我们在这里所阐述的,骨骼形成的过程也强烈地依赖于斑马鱼的负重。
一、斑马鱼概述
在基础和临床前研究中,斑马鱼已成为研究骨骼发育和疾病的重要模式生物。这些硬骨鱼的潜力在于它们的小体型,容易照顾,遗传顺从,和高再生能力。此外,由于胚胎和幼虫阶段的透明性,利用影响特定细胞或组织的转基因和突变株,可以更详细地监测骨生成和成骨细胞的活性。
这与胚胎、幼虫和成年个体长期体内成像的可行性相结合,从而将斑马鱼与其他脊椎动物模型(如啮齿动物)区分开来,在啮齿动物模型中,体内成像具有挑战性(表1)。
重要的是,斑马鱼的基因组包含约82%的人类疾病相关基因的同源基因,包括那些影响骨骼的基因。通过位点特异性重组酶(Cre)的组织特异性过表达和通过聚类规律间隔短回文重复序列(CRISPR)-Cas9的基因特异性敲除,以及反义寡核苷酸基因敲除方法在斑马鱼中都是可行的,这是进行正向和反向遗传筛选的理想条件。
此外,单核苷酸基因组编辑可以通过CRISPR-Cas9介导的敲除蛋白完成。药物可以通过多种方式给斑马鱼用药,包括直接将化学物质溶解在斑马鱼水/培养基中,这是药物筛选的方法(如下图表1)。

以上描述说明了斑马鱼研究骨骼生物学和疾病的潜力。斑马鱼作为骨骼研究模型的各个方面都得到了很好的评价。在这里,我们旨在通过介绍斑马鱼的骨骼结构和模式、细胞类型和基质矿化(依赖于负载)的基本信息,将斑马鱼介绍给更广泛的骨骼研究群体。
二、斑马鱼骨架的结构

图1所示 斑马鱼的骨架。(A)颅面和轴向骨骼的μ CT图像,包括由韦伯式的设备组成的脊柱、腹部(也称为前尾或胸)、尾、尾鳍椎骨。Hox基因表达模式显示。与哺乳动物骨骼相似,斑马鱼的肋骨与腹椎骨相连,保护内脏器官。(B)腹部和尾椎的近景。两个相邻的双锥形椎体(左)矢状面显示神经弓向背侧延伸并包围神经根。从腹椎骨向腹侧延伸的肋骨是铰接的,而尾椎骨延伸到包围尾动脉和静脉的血管弓。正面图(右)显示未矿化的椎体中心,包含脊索和空泡性软组织(未显示)。环状椎体终板区域由IVL连接(图中未显示),与斑马鱼的椎体生长区相对应。参数,包括VBL, V.Th和BV/TV,提供了有价值的措施来量化椎体的形态和结构。BV/TV =骨体积分数;IVL =椎间韧带;VBL =椎体长度;V.Th =椎厚度。
脊椎动物的骨骼一般分为外骨骼和内骨骼。斑马鱼骨骼的突出部分如下:(i)颅面骨骼,包括顶骨、颌骨和盖鳃的骨;(ii)轴向骨骼包括脊柱、肋骨、肌间骨以及非成对的背鳍、肛鳍和尾鳍(图1A).。斑马鱼的性成熟时间为90天,相应的标准长度(SL;从吻部到成年个体的后尾椎)为1.5到2.0厘米。斑马鱼不断生长,骨骼体积不断增大,体长可达3至4厘米,通常寿命为3年左右(也可达到5年)。
成年斑马鱼的头盖骨由74块颅面骨组成,远远多于哺乳动物的22块头骨。然而,斑马鱼的许多骨骼结构在哺乳动物中有明显的同源性,包括神经颅的前部,它类似于哺乳动物的上颚,以及在斑马鱼和哺乳动物之间保留的颅穹窿。和哺乳动物一样,斑马鱼头骨的特征是骨骼关节,包括纤维关节(如颅骨缝合)和下颌关节。
在脊柱形态方面,斑马鱼的椎骨数量与人类相似(斑马鱼为30 - 32节,人类为33节),生理曲度与人类相似:肋骨保护内脏的腹部区域脊柱后凸,尾端前凸。在颅尾目中,斑马鱼的脊椎由韦伯式的四节椎骨组成,连接着鱼鳔和耳朵(对声音的传递很重要),10个腹椎骨(也称为尾前椎骨或躯干椎骨)由杆状肋骨节连接,过渡到14个尾椎和3个尾鳍椎骨。与哺乳动物的椎管相似,脊髓通过从每个脊椎骨向背侧延伸的神经弓。尾椎也有向腹侧伸展并包围尾动脉和尾静脉的血管弓。
斑马鱼个体椎体中心的三维形态特征为其沙漏形状(图1B)与含有小梁骨和携带骨髓(BM)的哺乳动物脊椎骨不同,斑马鱼的脊椎体不容纳红色的BM,因为成年斑马鱼造血发生在肾脏(表1)。相反,它们充满了空泡状的脊索细胞,并被脂肪组织包围。在个体椎骨狭窄的中心周围发现了微米薄的小梁支撑。
虽然评估脊椎小梁骨(一种检测哺乳动物骨骼脆性的方法)在斑马鱼中是有限的,但椎体中心的形态通常被用作衡量脊椎骨骼质量的指标。3D的形态学参数,如椎体长度(VBL),骨体积(BV)、骨体积/组织体积(BV /TV),椎横断面厚度(V.Th)和偏心率(圆度)提取微计算机断层扫描(μCT)扫描的斑马鱼脊柱(图1B),允许定量的变化,例如,由于肌肉骨骼活动改变,衰老和疾病。
此外,血管弓和神经弓的厚度和体积用于评估斑马鱼突变体的椎体形态。小型斑马鱼的一个优点是可以在高分辨率下分析完整的骨骼(例如,全身的μ CT扫描,像素大小为1 μm2),比在大型啮齿动物物种中更快。这提供了一种可能性,既可以评估整个机体的整体三维形态,又可以同时在高分辨率下表征组织形态,这已在变形的骨关节炎椎体中完成。此外,纵向组织学切片或整片染色,如茜素红染色,可以在显微镜分辨率下显示完整的骨骼。

图2所示 斑马鱼骨基质成分的评估。(A)拉曼光谱可以绘制再生尾鳍骨(此处为7 dpa处的矿物与基质比率)近端和远端截肢平面(箭头)的矿物相关和蛋白质相关特性。再生部位的拉曼光谱(灰色曲线),蛋白质相关的峰包括酰胺I、酰胺III和羟脯氨酸,与矿物相关的磷酸盐峰相比更显著,反映出与天然骨组织相比更低的矿物-基质比率(黑色曲线)。(B) qBEI可用于评估斑马鱼骨骼中的矿物密度分布(此处为脊柱矢状面和终板区域特写)。由感兴趣区域内的校准像素强度生成的直方图允许提取Camean、Capeak和Cawidth,以及具有Calow和Cahigh的区域。Cahigh=高矿化程度;Calow=低矿化程度;Camean= Ca均值含量;Capeak= Ca峰含量;Cawidth= Ca分布的异质性;dpa =截肢后天数;qBEI =定量背散射电子显微镜。
和哺乳动物一样,斑马鱼的脊椎是由软组织连接在一起的,这有利于运动,也增加了运动的范围。
在哺乳动物中,椎间盘(IVD)由纤维软骨软骨(纤维环)和髓核组成。相比之下,斑马鱼的椎间软组织的特征是环状韧带(椎间韧带[IVL])连接相邻两个椎体的外层圆形边缘。虽然IVD在人体中的生理作用是阻尼重力载荷的压缩力,但斑马鱼的脊柱是轴向加载的,这是由于在粘性的水中游泳产生的压缩力和附着在椎骨上的肌腱直接传递的肌肉力。与重力垂直作用于脊柱的四足动物(如老鼠、狗)相比,斑马鱼在负重方面更具优势。
因此,它们为研究脊柱和椎间组织退行性变以及运动模式改变对骨-肌腱-肌肉单元的影响提供了一种有价值的工具,这可以通过组织学和先进的x线成像方法在小型斑马鱼中进行评估(图2)。
三、斑马鱼轴向和鳍骨架的图案
和其他脊椎动物一样,斑马鱼骨骼的轴向模式表现在脊柱上,脊柱沿着前后轴被划分为不同类型的脊椎。对小鼠和鸡的研究揭示了不同类型脊椎的形态多样性和轴向位置,这些脊椎对区域Hox基因表达编码的位置线索敏感。
虽然人类中的HOX突变导致早期发育致死,但已经描述了与HOX相关的轴向骨骼缺陷,以及四肢和颅面缺陷、关节炎和各种类型的癌症。斑马鱼轴向骨骼的分区受到相同的机械控制,依赖于空间(和时间)共线性;即染色体上基因的物理序列与其表达前后边界(和时间)之间的对应关系(图1A)。
哺乳动物基因组中有39个Hox基因分布在4个集群中,而斑马鱼有48个基因分布在7个集群中。这些差异是脊椎动物进化过程中基因组复制和基因丢失的结果。
因此,Hox表达域和轴向结构区划性在斑马鱼和四足动物之间只有部分相同(如Hoxc6和Hoxd12,但Hox9不相同)。然而,研究胚胎和幼体斑马鱼的同源性转化可能是揭示Hox蛋白多种调节功能的有用工具。
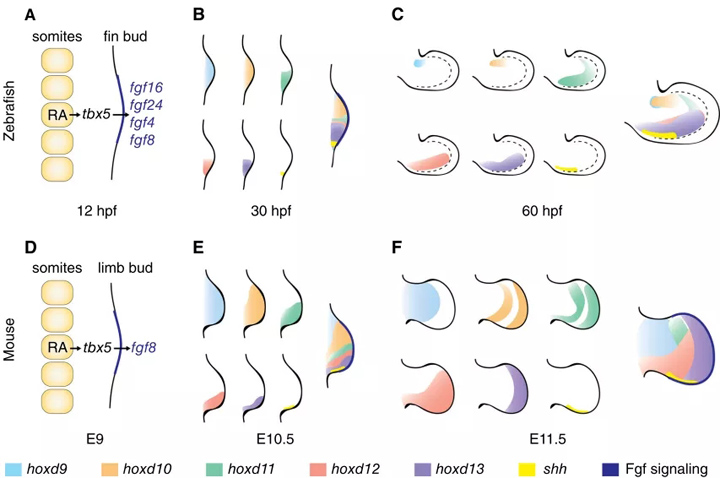
图3所示 四肢和胸鳍芽的早期模式是相似的。(A - C)斑马鱼鳍芽的早期模式。(A)在12 hpf时的鳍诱导:来自孢子虫的RA激活信号,导致侧板tbx5和远端芽Fgf信号的激活,建立AER。(B) Hox基因和shh在30hpf的早期表达。(C) Hox基因和shh在60hpf的表达。需要注意的是,在30 hpf和60 hpf处,后翅芽区的hoxd11表达域仍然局限于该区域。虚线=在芽适当和鳍折边界。(D)类似于A中描述的鳍芽诱导信号在小鼠E9位点出现。(E) Hox基因和shh在E10.5小鼠肢体中的早期表达与在鳍芽中的B中观察到的模式相似。hoxd9和hoxd10贯穿整个花蕾,而hoxd11、hoxd12、hoxd13和shh则局限于后端区域。(F) E12.5小鼠四肢Hox表达域与鳍芽C中Hox表达域的差异;hoxd11、hoxd12和hoxd13域向更前面延伸;嘘的声音仅限于臀部。从左到右=近端远端轴,从上到下=前后轴。E10.5 =胚胎期10.5天;E12.5 =胚胎期12.5天;E9 =胚胎第9天;hpf =受精后小时数。
在鱼类中,轴向骨骼包括脊柱和相关的中鳍(背、肛和尾),而成对的胸鳍和骨盆鳍位于腹部区域的腹外侧。斑马鱼的背鳍和臀鳍的位置尚未被研究过,但推测可能与鲨鱼一样,是由Hox和Tbx基因的表达域决定的。
在Hox表达和维甲酸(RA)等诱导信号方面,胸鳍的发育与四足动物四肢的发育表现出高度的相似性,特别是在早期阶段(图3A-F)。在原肠形成过程中,RA负责边缘区定位。
随后,RA在体节中产生并激活中间中胚层的Wnt2b,而Wnt2b又激活侧板中胚层的Tbx5 。Tbx5是前肢启动过程中较早的转录因子,然后触发成纤维细胞生长因子(Fgf)信号,建立转录级联,调节肢体近端和远端模式。
在四足动物和鱼,两个信号附体的组织者在原基形成:顶端外胚层的脊(AER)远端芽,调节肢体和鳍产物通过Fgf信号和极化区活动(ZPA),后声波刺猬的表达(嘘)和调节前后的模式,非脊外表皮指示背腹模式。
虽然这些机制显然是保守的,但鳍和肢在后期的调控和模式差异表明,表达模式和信号机制在进化过程中已经被修改,即在鳍到肢的过渡过程中,这可能导致了新骨骼特征的产生,比如四足动物的趾。在四足动物和鱼类(包括斑马鱼)中对Hox基因表达和调控这些和其他模式决定因素进行对比,将在未来揭示骨骼进化和人类先天模式缺陷的更多方面。
四、骨化类型和骨细胞
在斑马鱼中,受精后3—4天,骨骼开始发育,并在2 - 4个月时逐渐形成成熟的骨骼(取决于斑马鱼的大小,表1)。许多元素通过膜内和软软骨骨化形成,还有一些元素通过软骨内骨化形成(表1)。
斑马鱼骨骼根据解剖区域的不同,是由三种骨化类型中的一种形成的。膜内骨化是斑马鱼骨骼骨化的主要形式,主要发生在颅顶、覆盖鳃的鳃盖和大部分椎骨等部位。软骨内骨化,即软骨模板依次被骨取代,只产生包括1至5椎体神经弓在内的少数元素。软软骨骨化发生在软骨表面而不取代软骨模板,发生在斑马鱼下颚。虽然哺乳动物的椎骨完全通过软骨中间形成,但斑马鱼中心直接矿化从脊索鞘,脊索周围的骨化层,然后膜内骨形成。值得注意的是,脊索鞘也负责在中央形成之前斑马鱼脊柱的分割。
基于脊椎动物骨骼基因的进化保守性,斑马鱼骨骼中含有与哺乳动物相同的骨细胞,即成骨细胞、破骨细胞和骨细胞(表1)。
与哺乳动物一样,骨结合是由骨祖细胞衍生的成骨细胞完成的。在斑马鱼骨骼中,骨表面有很大一部分被成骨细胞覆盖,成骨细胞的形态变化很大,并依赖于其位置和功能。在脊柱中,成骨细胞骨形成导致椎体前后方向体积和长度增加,从而拉长脊柱。在椎体间生长区域,特别是在圆形椎体终板上,可以很容易地评估这种骨形成(图1B)。通过使用静态组织形态测定法(静态组织形态测定法通常应用于啮齿动物骨骼或人体活检),可以分析斑马鱼脊柱中的骨骼,包括每骨周长成骨细胞数量(N. Ob/B.Pm)、每骨表面类骨质表面(OS/BS)和类骨质厚度(O.Th)。
然而,斑马鱼骨也可以在连续的时间点用钙黄素或素红等荧光染料标记,提供了进行动态组织学形态测定的机会,并确定骨形成率(BFR)和骨矿物附着率(MAR)(表1)。在啮齿类动物或人类模型中,染料是通过注射或摄入来使用的,而斑马鱼通常是通过将它们浸泡在染料溶液中来染色的。
斑马鱼的骨吸收是由单核和多核破骨细胞完成的。尽管单核破骨细胞存在于发育的早期阶段,并且与浅层骨吸收模式有关,但多核破骨细胞会产生骨吸收陷窝,这在哺乳动物后来的破骨细胞中很常见。
哺乳动物骨骼建模和重塑的概念也适用于斑马鱼。骨造型被定义为通过骨形成和骨吸收来适应不同表面载荷增加或减少的过程,而重塑则是在同一位置进行,以维持骨基质质量和修复微裂纹。重塑,包括机械敏感骨细胞对破骨细胞和成骨细胞的协调,以更新骨和修复微裂纹,在斑马鱼中不太明显。骨基本上不存在。然而,斑马鱼的重塑过程被认为是位点依赖性的,并与终生生长的需求有关。
斑马鱼的骨头,与medakfish (Oryzias latipes,另一种硬骨鱼)的骨头相反,通常是骨细胞性的,尽管脊椎骨在幼年早期不包含骨细胞,而骨鳍射线和鳞片在整个生命过程中都保持骨细胞性。尽管哺乳动物骨细胞是重塑的主要协调者,但只有少数研究关注斑马鱼骨细胞腔隙网络和树突特征的机械感测能力。
然而,有证据表明在斑马鱼OI模型中骨细胞网络的形态与骨形成之间存在关系,骨形成的改变与骨细胞陷窝数量的急剧减少有关。这些信息揭示了斑马鱼和哺乳动物骨骼生物学之间的一些重要差异,但也有许多共同之处(表1)。
五、斑马鱼的骨基质和矿化
脊椎动物的骨是由一种主要含有I型胶原蛋白和非胶原蛋白的软基质组成,并通过磷酸钙骨的掺入而变硬。
虽然不同脊椎动物骨骼的矿化程度不同,但哺乳动物和斑马鱼骨骼基质的基本大分子和元素组成是保守的。用振动光谱学方法证明了斑马鱼骨中存在磷酸盐、碳酸盐、酰胺I和酰胺III、脯氨酸、羟脯氨酸和苯丙氨酸。这些分析显示了嵌有碳酸盐骨磷灰石的胶原基质的典型指纹。
在鱼骨生长过程中,无定形磷酸钙被认为在组织成熟过程中转变为更结晶的矿物。除了钙(Ca)和磷(P),镁也是储存在斑马鱼骨头中的主要矿物质之一。此外,包括锶和锌在内的微量元素参与了骨骼的形成,这与哺乳动物的情况类似。特别是,这些元素分别在scale和椎骨基质中使用x射线荧光显微镜和能量色散x射线光谱学检测到。
显然,硬骨动物和陆生哺乳动物在矿物质代谢方面存在一些差异。四足动物依赖膳食中磷和钙的摄入来维持钙磷基骨基质,而斑马鱼生活在富含钙的环境中,并通过鳃吸收钙。然而,膳食中磷的摄入是必需的,通过基因控制或减少饮食来降低磷水平会导致非矿化基质。
和哺乳动物一样,斑马鱼骨骼基质的组成和组织对于提供抗骨折能力至关重要。具体来说,相关的胶原蛋白和矿物质和相关结构属性(胶原蛋白对齐、酶和非酶的交联,矿物粒子的大小和方向)和成分属性(碳酸盐磷酸盐比),以及矿化的程度而言,整体骨矿物质密度(BMD)或当地Ca内容和分布,确定组织的机械性能水平。
这意味着整个骨骼水平的骨折风险。从哺乳动物骨骼分析中借鉴了几种技术来确定微米大小的斑马鱼骨骼的矿化程度(表1)。基于羟基磷灰石幻影校正的μ CT已被用于评估斑马鱼脊柱的骨密度。的确,突变导致的骨密度变化支持斑马鱼和哺乳动物模型中存在类似的矿化途径。通过振动光谱学分析了携带col1a1突变的斑马鱼病理逻辑骨基质的大分子组成,并证实了矿物-基质和碳酸盐-磷酸盐比例的改变。
此外,可以使用振动光谱监测截肢后再生的斑马鱼鳍骨的再矿化,从而比较新骨质量和天然骨质量(图2A)。钙含量和钙在骨基质中的分布是骨基质矿化良好的另一个指标。这些参数通常在啮齿动物和人类活检中使用二维(2D)反向散射电子显微镜(qBEI)进行评估(图2B)。近来,qBEI也被用于评价斑马鱼的脊椎,平均钙含量增加8%与肌肉骨骼活动增加有关。
矿化的增加可能会增加骨骼的抗骨折性,尽管对斑马鱼骨骼(如单个椎骨)进行全骨力学测试是具有挑战性的(尽管可能),因为它们的尺寸较小(VBL = 500 μ m)。纳米压痕技术是在组织水平上评估斑马鱼椎骨机械性能的有价值的工具。例如,衰老过程中,Ca/P比值的升高与斑马鱼脊椎弹性模量的增加有关,然而,也可以观察到相反的效果。
在胶原紊乱和连续的矿物颗粒沉积的情况下,椎骨的弹性模量会降低,尽管矿化程度较高,这就强调了组织良好的基质矿化对于抵抗骨折的重要性。值得注意的是,斑马鱼的骨基质弹性模量值与哺乳动物相似(高达24 GPa),突出了两者在骨基质组成和力学特性方面的相似性。这证实了使用斑马鱼是一个研究遗传改变和外部刺激对骨基质质量影响的模型。
基质矿化的问题,如磷酸酯内稳态紊乱,已在斑马鱼中出现。磷酸盐是骨中羟基磷灰石形成的必要矿物质,其缺乏(低磷酸盐血症)是在突变体中引起的,在突变体中,磷酸盐调节的必要基因发生改变。无骨(头)突变体ectonucleoside triphosphate diphosphohydrolase 5 (entpd5)基因受到影响(entpd5),没有矿化骨形成。在另一个突变体,dragonfish (dgf),ectonucleotide pyrophosphatase 1 (enpp1)基因突变,导致焦磷酸盐水平降低和轴骨和颅面骨骼的异位矿化。
dgf变异显示改变与磷酸自我调节有关的基因的表达和骨矿化,如fgf23、溶质载体家族34个成员1 (slc34a1a,也称为npt2a) entpd5,分泌phospho-protein 1 (spp1),也可以被用来模型广义动脉钙化的婴儿期(GACI)和弹性纤维假黄瘤 (PXE)。在双突变体nob/dgf斑马鱼中观察到恢复的骨骼矿化,表明Entpd5和Enpp1的相互调节。这说明了斑马鱼突变体分析和全新技术在理解脊椎动物矿化缺陷方面的有用性。
六、斑马鱼骨架对载荷的响应
尽管斑马鱼骨骼的负重不同于陆生动物,但由于水的支持性浮力,骨骼确实会对机械载荷做出反应(表1)。这来自于骨骼肌肉收缩的作用,以及在粘性介质中游泳时产生的反作用力。

在胎儿早期,所有脊椎动物都是在水环境中发育的,无论是在子宫内、卵中还是水中(表1)。在这段时间内,肌肉对骨骼组织的作用引起了对骨骼发育的生物力学刺激。在小鼠、小鸡和人类的实验中已经证实,胎儿运动的限制会导致矿化改变和关节突起形态形成的失败。
在斑马鱼中,基因或药理学诱导的麻痹导致软骨细胞成熟改变,并通过改变软骨细胞增殖和迁移导致异常关节形态发生。有限元(FE)建模的幼虫颚,其中的结构被细分为更小和更简单的实体,允许建模的负荷影响组织变形,并已证明改变关节形状影响生物力学应变的模式。
众所周知,关节的生物力学负荷是骨关节炎的关键危险因素。此外,像col11a2和prg4这样表现出关节形状改变的突变体会在这些关节中发展成骨关节炎。鉴于近来相关的识别位点与骨关节炎和关节的形状改变人类发生前症状和功能性筛选平台形式的需要,这就提出了一个利用斑马鱼的前景筛选骨关节炎易感基因与联合开发和维护(图4)。
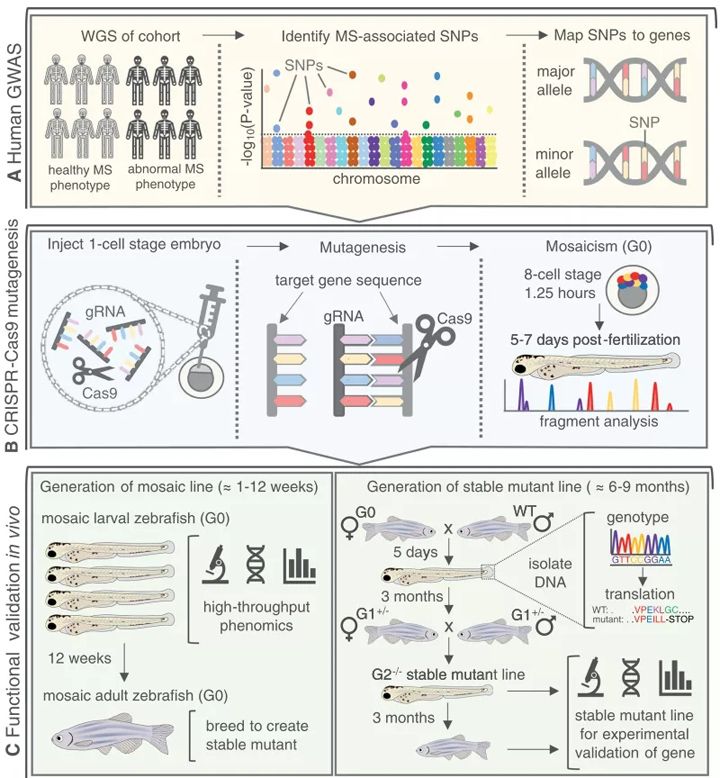
图4所示 斑马鱼作为快速验证gwas衍生的MS疾病相关候选基因的工具。(A)人类GWAS在大队列上进行WGS,并在异常MS表型的人群中识别SNP突变。然后利用全外显子组测序将snp映射到近期的基因上。(B) CRISPR-Cas9技术可用于体内靶向敲除诱变。设计基因特异性gRNAs,并将Cas9注入到单细胞阶段的斑马鱼胚胎中。高效的gRNAs会促进目标基因外显子内的双链断裂,导致indel突变。一个花叶(G0)胚胎(crispant)将发育,包含各种突变和WT等位基因的兴趣基因,并通过片段分析验证。(C)(左)马赛克斑马鱼幼虫在骨骼发生过程中可以快速筛选MS表型,为基因验证提供了一种高通量和高效的表型组学方法。花叶G0斑马鱼可以被饲养到成年,用一个已知的单一突变等位基因繁殖成一个稳定的系。(右)G0花叶斑马鱼与WT杂交,导致杂合幼虫(G1 +/−)夹鳍分离DNA进行基因分型,并通过硅转译识别导致过早终止密码子的等位基因,损害蛋白质。具有相同突变等位基因的G1 +/−相互繁殖,产生具有稳定突变的相关基因的G2−/−系。稳定的突变体幼虫或成虫可用于MS相关表型的实验验证。gRNA =引导RNA;hpf =受精后的小时数;女士=肌肉骨骼;单核苷酸多态性;全基因组测序;WT =野生型。
斑马鱼的脊柱和鳍的矿化比运动开始的时间要晚得多,在大约7 dpf (SL = 3.8 mm)时首次观察到椎体的矿化,在第14天左右观察到椎弓和鳍条(SL = 5 - 6 mm)。成年斑马鱼的骨骼肌生理负荷增加可触发椎骨形成,这表明斑马鱼的骨骼易受Wolf定律的正向骨建模影响。斑马鱼每天在5-14 dpf范围内进行9小时的游泳训练后,其尾部和脊柱出现过早骨化。
陆生物种中,骨细胞具有机械传感器的功能,通过调节糖蛋白硬化蛋白(SOST),指导成骨细胞和破骨细胞的重塑活动。然而,骨细胞鱼也模仿骨头对负荷的反应。对骨细胞斑马鱼和骨细胞青鳉进行游泳训练后,两种鱼的新骨形成模式惊人地相似,这是由软骨细胞和成骨细胞在单个椎骨高应变区域的sost表达介导的。有趣的是,斑马鱼的椎体运动分析和FE可以预测负重时骨衰竭的模式,这可以用于测试突变体的骨性能。
重力对陆生动物骨骼的负重起着重要作用,长时间暴露在微重力(失重或重力接近于零)下会导致人类骨密度下降。令人惊讶的是,重力也会影响硬骨鱼的骨骼。利用国际空间站(ISS)上的一个水生生境进行了几项关于青鳉鱼的研究。
这些研究表明,暴露在微重力环境下,青鳉会失去骨密度,骨骼基因转录发生变化,破骨细胞活性增加。虽然在国际空间站上没有饲养过斑马鱼,但它们已经暴露在越来越大的重力(超重力)下。在斑马鱼幼虫发育过程中暴露于3g至9g可改变软骨细胞成熟,并改变骨骼基因的矿化和转录。这些研究表明,斑马鱼模型在研究负重对骨骼的影响方面具有多功能性。
七、结论
斑马鱼在骨骼疾病和再生研究领域的应用越来越多,因为它们易于遗传操作,方便的药物治疗选择,以及在体内成像的可能性。高通量的遗传和药物筛选可以在斑马鱼幼虫中进行,也可以用于异种移植受体研究骨转移。哺乳动物和斑马鱼的生理机能有许多相似之处,这使得研究斑马鱼的激素性骨病成为可能。
参考资料:
[1] Spoorendonk KM, Hammond CL, Huitema LFA, Vanoevelen J,Schulte-Merker S. Zebrafifish as a unique model system in boneresearch: the power of genetics and in vivo imaging. J Appl Ichthyol.2010;26(2):219-224.
[2] Busse B, Galloway JL, Gray RS, Harris MP, Kwon RY. Zebrafifish: anemerging model for orthopedic research. J Orthop Res. 2020;38(5):925-936.
[3] Xu C, Volkery S, Siekmann AF. Intubation-based anesthesia for longterm time-lapse imaging of adult zebrafifish. Nat Protoc. 2015;10(12):2064-2073.
