
 服务热线:0571-83782130服务热线:0571-83782130
服务热线:0571-83782130服务热线:0571-83782130
NEWS
新闻中心
SERVERS
服务项目
NEWS
最新资讯
发布时间:2021-06-01
核心摘要
造血是一个复杂的过程,不成熟的骨髓前体细胞成熟为各种类型的血细胞。虽然造血系偏向(包括贫血和中性粒细胞)与慢性炎性疾病的关系早已被人们所认知,但其原因尚不清楚。
近来,在斑马鱼模型中,细胞质多蛋白炎性小体复合体被证明可以激活炎症和免疫反应,并直接调节造血功能;这被认为是通过裂解和失活红细胞转录因子GATA1而发生的。本文总结了目前可用的斑马鱼模型,用于研究炎性小体介导的造血调节这一未被重视的作用,并提出了用于治疗与人类慢性炎症性疾病相关的造血改变这一新的治疗策略。

一、慢性炎症中的造血系统偏向
造血是发生在胚胎发育和成年期的血细胞形成的完整过程。在脊椎动物中,这个过程包括两个阶段。
首先,在胚胎发育过程中,原始造血波(见术语表)是短暂的,红细胞和髓系祖细胞缺乏多能性。在第二阶段的造血波,发生在发育的后期阶段,造血干细胞(HSCs)出现,并在成年生物体中产生各种类型的血细胞。
HSC的成熟包括淋巴细胞(T细胞,B细胞和自然杀伤细胞(NK))、骨髓细胞(粒细胞和巨噬细胞)和红细胞(巨核细胞,红细胞)的分化。多个信号通路通过不同的转录因子调控造血细胞的成熟、增殖和分化来协调这一复杂的过程。转录因子GATA1和SPI1(又称PU.1)是必不可少的,并表现出交叉抑制关系(图1):GATA1调节红细胞分化,SPI1调节骨髓细胞分化。

图1所示。斑马鱼和小鼠中典型炎性小体对HSPC谱系决定选择的调控。HSPCs中炎症小体的激活导致GATA1的分裂和失活,促进促进髓细胞分化的SPI1数量的增加。相反,抑制HSPCs中的炎性小体导致GATA1量的增加,从而促进红细胞分化。简称:CASP1, caspase 1;造血干细胞,造血干细胞和祖细胞。
然而,考虑到许多未知的途径似乎在这一复杂的生物学过程中发挥了作用,关于可能导致终端红细胞和骨髓细胞分化的因素以及HSC生态位在这些过程中的参与仍存在许多争议。
例如,当造血前体中的GATA1直接作用于红系-巨核细胞系统时,红系细胞中GATA1的过度表达抑制了它们在小鼠中的终端分化。
此外,GATA1的表达和活性在红细胞分化初期增加,但在后期均降低,从而使小鼠和人类的红细胞分化达到终点。所有这些复杂的途径都可能具有重要的临床意义,因为造血系统偏向(主要是中性粒细胞减少症和贫血)与疾病的发生率增加有关,至少包括30种具有明显炎症成分的病理,包括感染、类风湿性关节炎、动脉粥样硬化、炎症性肠病、肾脏和肺部疾病、自身免疫性疾病、神经退行性疾病和癌症等。
在这篇综述中,总结了哺乳动物炎性小体的一般结构和功能,以及巨噬细胞和嗜中性白细胞炎性小体的特点。接下来是关于使用斑马鱼对炎症和炎症小体进行建模的主要优势的讨论,描述了该模型如何突出炎性小体在造血中的重要性。确实,该模型可能有助于解释与人类慢性炎症相关的造血系统偏向,从而揭示了新的候选治疗靶标,可用于今后对其中某些疾病的研究。
炎性小体作为脊椎动物固有免疫系统的传感器,在各种刺激下促进炎症反应,包括感染、组织损伤或代谢失调。它们是由受体,包含CARD(ASC)的凋亡相关的斑点状蛋白和通过同型相互作用的效应子形成的多蛋白复合物。
近来,发现鸟苷酸结合蛋白(GBP)家族在人类、小鼠和斑马鱼中也形成了这些多蛋白复合物的一部分。炎性小体通过激活炎性半胱天冬酶发挥作用,即半胱天冬酶1(CASP1)(典型炎性小体)和CASP4、CASP5(人类)和Casp11(小鼠)(非典型炎症小体)。炎性小体传感器根据其结构分为核苷酸结合域(NOD)样受体(NLRs),在黑色素瘤2(AIM2)样受体(ALRs)或PYRIN中缺失。当检测到特定的刺激,与传感器相互作用并促进ASC的低聚,形成斑点,并能聚集并激活炎性半胱天冬酶。
与FMF和CAPS相关的贫血常见的地中海热(FMF)和冷卟啉相关周期性综合征(CAPS)是由编码炎性小体成分的基因的种系突变驱动的两种炎症性疾病。FMF是全世界常见的单基因自身炎症性疾病,约有15万名患者受到影响。超过60%的FMF患者出现贫血,贫血与铁状态有关,而不是与炎症有关,以血浆IL-6浓度测定。
同样,抗铁性低色素性贫血也与CAPS有关。HSPCs中炎性小体的过度活化可通过促进红细胞生成来直接促进贫血,这解释了这些疾病通常的铁难治性,但这仍有待进行深入研究。
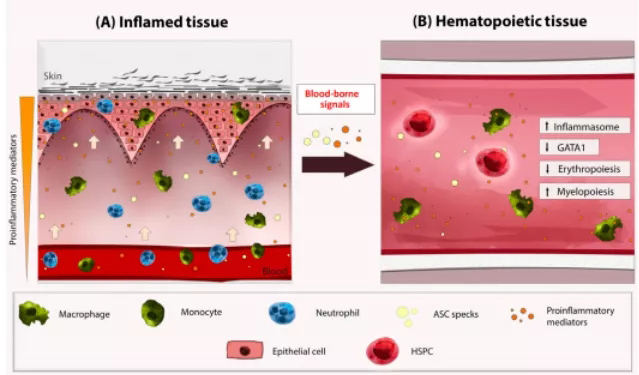
此外,释放的ASC斑点能够在CAPS小鼠模型中传播炎症,并且能在CAPS患者的血清中发现,但在FMF中没有发现。因此,巨噬细胞衍生的ASC斑点可能被HSPCs吸收,激活其炎性小体,以红细胞生成为代价进一步增强骨髓生成,这可能解释了与CAPS相关的耐铁性贫血(参见临床医生角和悬而未决的问题)(图2)。

图2.(A)在这个模型中,主要基于斑马鱼的研究,巨噬细胞和中性粒细胞被招募到发炎的组织中,例如皮肤,在那里它们被不同的促炎信号激活。炎性小体的激活导致ASC斑点释放到循环中。(B)血源斑点可以到达造血组织(哺乳动物的骨髓和斑马鱼的肾骨髓),被HSPCs或巨噬细胞吸收,激活其炎性小体,导致IL-1β/IL-18释放,GATA1分裂,红细胞生成减少,骨髓生成增加。缩略语:ASC,凋亡相关斑点样蛋白,含CARD;造血干细胞,造血干细胞和祖细胞;IL,白介素。
长期以来,CASP1的自身蛋白分解被认为是由其活性亚基p20和p10的邻近性触发的;然而,这一观点近来受到了质疑。作者在小鼠模型中证明,CASP1活性二聚体的优势种实际上是全长p46二聚体和一个瞬时种p33/p10。
此外,p33/p10在炎性小体大小和细胞类型指定的动力学下发生了自动处理,导致p20/p10物种的释放变得不稳定,蛋白酶活性终止。因此,炎症小体也包含了内在的自我限制机制,确保CASP1及时失活。
Caspase二聚和自身蛋白水解成熟导致促炎细胞因子白细胞介素-1β (IL-1β)和IL-18[25]的加工和分泌,并诱导一种称为焦亡[18]的程序性细胞死亡形式。细胞焦亡导致膜完整性的丧失,进而介导细胞内内容物释放到细胞外液。IL-1β/18的释放和焦亡都是由Gasdermin D(GSDMD)介导的。
除了在感染和炎症中的作用外,炎性小体在维持基本细胞功能方面也很重要,如自噬、代谢、细胞增殖、基因转录和肿瘤发生。近来的研究结果也表明炎性小体在斑马鱼、小鼠和人类的造血调节中起着重要作用。这是相关的,因为不受控制的炎性小体激活导致了某些人类自身免疫和代谢紊乱,强调了在生理和病理背景下更好地理解这一过程的重要性。
三、细胞型特异性炎性小体
中性粒细胞和单核细胞/巨噬细胞属于髓系,起源于一种常见的粒细胞-单核细胞祖细胞(GMP),来源于造血干细胞的分化。
中性粒细胞是大多数哺乳动物中丰富的循环白细胞(50–70%)来自骨髓(BM)的CXCL8、CXCL1和CXCL2趋化因子信号。以类似的方式,单核细胞在CCL2和CCL7信号通过CCR2从BM中释放。随后,它们被炎症组织附近激活的内皮发出的信号渗出[38–40]。在被称为“危险动员”或“紧急造血”的过程中,中性粒细胞的产生在感染或炎症期间增加,主要由造血细胞因子粒细胞集落刺激因子(G-CSF)触发。
中性粒细胞和巨噬细胞炎性小体通过典型和非典型途径感知感染和无菌性炎症。这两种细胞类型主要表达广泛的NLR和ALRs,是很早参与炎症过程的细胞(表1)。
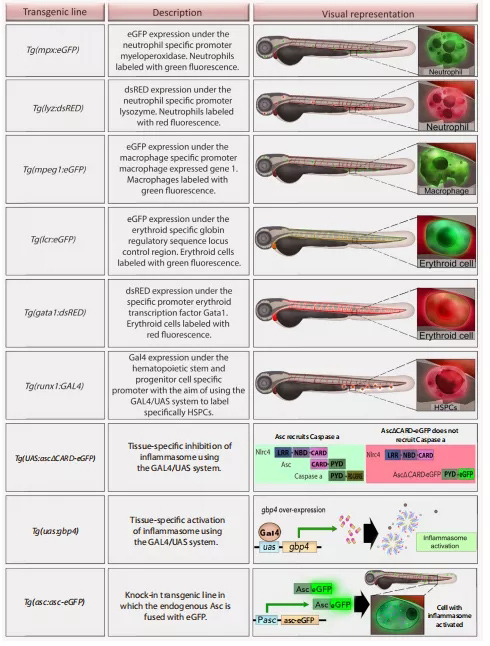
表1. HSPCs、红系祖细胞和成熟髓系细胞中主要典型炎性小体的组成和效应功能(巨噬细胞和中性粒细胞)a,b
尽管如此,这些细胞中的炎性小体之间存在差异,从而能够复杂而特异地调控细胞过程。尽管它们的NOD-、LRR-和吡啉结构域含蛋白3 (NLRP3)和NOD-、LRR-和CARD结构域含蛋白4 (NLRC4)的表达量相似,但NLRP12在小鼠中性粒细胞中的表达量高于巨噬细胞。人巨噬细胞中NLRC4的激活是通过特定的革兰氏阴性细菌成分进行的。
巨噬细胞通过非ASC依赖的NLRC4信号或ASC依赖的NLRP3和AIM2炎性小体激活诱导焦亡,促进细胞内病原体的清除。尽管NLRC4或AIM2信号也在人和小鼠中性粒细胞中被观察到,但它们似乎并不会导致焦亡。因此,嗜中性粒细胞对细菌的焦亡和细胞内感染具有相当的抵抗力。
因此,它们也参与IL-1ß的持续释放,从而扩增中性粒细胞的招募和激活。
此外,当人类中性粒细胞炎症小体被典型激动剂激活时,IL-1ß释放,而不是IL-1α和IL-18,表明IL-1β和IL-18蛋白的释放在人类中性粒细胞中有差异调控。
NLRP3炎症小体参与炎症和代谢紊乱以及宿主防御。虽然,它也存在于中性粒细胞中,但它的作用不同。中性粒细胞中的NLPR3信号通路只能通过可溶性激动剂激活,但它可以被小鼠巨噬细胞中的可溶性和颗粒状或晶体状激动剂触发。
此外,NLRP3激活在巨噬细胞和中性粒细胞中导致不同的细胞结果。NLRP3/ casp1依赖和NLRP3/ casp8依赖的炎症小体均可导致小鼠巨噬细胞的焦亡和凋亡,而中性粒细胞则不能。
关于造血干细胞和祖细胞(HSPCs),炎性小体的表达/激活很少受到关注。然而,NLRP3和NLRP1炎性小体的主要组成部分的编码基因,包括NLRP3、ASC和CASP1,在人和小鼠HSPCs中都有表达。同样,斑马鱼胚胎中新生的HSPCs也被报道表达ASC。因此,每种细胞类型显示不同的炎性小体成分、激活机制和效应功能。
四、疾病中的炎性小体
来自小鼠模型的大量数据及人类临床数据的支持,强烈支持炎性小体在疾病的起始和/或进展中的作用,并对公共卫生产生强烈影响;这些包括某些代谢紊乱和神经退行性疾病。
然而,支持炎症小体在人类病理学中起关键作用的强有力的证据来自各种疾病的众多遗传关联研究,与它们在炎症和组织修复调节中的重要性一致。
令人信服的证据来自炎症小体传感器MEFV (PYRIN编码)、NLRP3和NLRP1编码氨基酸的点突变,它们分别导致家族性地中海热(FMF)、冷PYRIN相关周期性综合征(CAPS)以及皮肤炎症和癌症易感综合征。凸显了这些蛋白的临床意义。
此外,近来的两项研究表明,慢性炎症性疾病(如慢性阻塞性肺疾病(COPD)和慢性阻塞性肺疾病(CAPS))患者的血清中存在ASC斑点,这些斑点可能被认为是活动性疾病的生物标志物。据推测,细胞外斑点可能也有助于扩大炎症,以及嗜中性粒细胞和贫血的人类慢性炎症疾病,如这些;他们可能通过改变HSPCs的红髓系命运来达到这一目的,但这仍有待证实(图2)。
从另一个角度来看,动脉粥样硬化斑块中中性粒细胞的存在与这些斑块的不稳定性和动脉粥样硬化的不良预后有关,这种不稳定性也与中性粒细胞胞捕获网(NETs)的形成和炎性小体的激活有关,因为NETs已经在人和小鼠的动脉粥样硬化病变和动脉血栓中检测到,并且可以诱导内皮细胞、抗原提呈细胞和血小板的激活。
此外,两种类型的细胞死亡,即焦亡和网状增生,都需要GSDMD的切割;然而,小鼠中性粒细胞利用炎性小体依赖机制来切割GSDMD并激活网状增生,人类中性粒细胞却利用中性粒细胞弹性蛋白酶来实现这一点。
总之,这些观察表明,慢性激活的巨噬细胞和中性粒细胞炎性小体可以破坏体内平衡,并可能导致炎症性疾病;因此,炎性小体可能是在各种病理中恢复体内平衡的有希望的治疗靶点,但这显然仍有待进一步研究。
五、斑马鱼:研究造血和炎症的独特模型
斑马鱼已经成为许多生物医学研究领域的特殊模式生物。
它是唯一一种具有无脊椎动物(例如果蝇和线虫)主要优势的脊椎动物模型,包括:
(1)繁殖力高(每只雌虫每周可获得200 - 300个卵),胚胎发育迅速且外化;
(2)胚胎是透明的,可以利用光学和荧光方法在组织和器官中发育和追踪其细胞;
(3)该动物适合进行高通量化学和遗传筛选。
此外,其基因组已被完全测序,其70%的基因与人类相同,其中高达82%的基因已知与人类疾病相关/致病。这种高度的遗传同源性允许转基因研究与人类基因,并在体内研究基因调控和功能。
斑马鱼被用作研究造血的研究模型,虽然斑马鱼的造血功能发生在不同于其他脊椎动物的部位(例如,斑马鱼造血干细胞驻留在肾骨髓中),但控制造血功能的遗传程序是保守的。
斑马鱼与包括人类在内的哺乳动物之间的关系,使斑马鱼成为临床相关模型系统。前面提到的优点,加上详细的基因组数据库的构建,使得该模型可以使用一系列基因方法来研究各种血液疾病,如先天性和获得性贫血、中性粒细胞和白血病。斑马鱼在免疫、炎症和感染领域也巩固了其作为强大动物模型的地位。它有助于提高我们对人类感染的认识,例如很难在哺乳动物体内建模的结核病。
此外,斑马鱼早期生命阶段的光学透明性为研究体内炎症小体的激活和动力学提供了前所未有的可能性。
胚胎中获得(mRNA)和功能丧失实验[morpholinos,转录激活因子样效应核酸酶(TALEN)和CRISPR-Cas技术]相对简单,可以研究特定分子的作用,包括参与炎症小体的分子,利用不同的刺激物和病原体可以更好地解释这些作用。
此外,斑马鱼还可以分析和可视化不同类型的免疫细胞,包括中性粒细胞和巨噬细胞;事实上,炎症小体的组装已经通过转基因斑马鱼系进行了研究,这些斑马鱼系要么已经可用,要么容易产生。在众多可用于研究造血和炎症小体的转基因系中,需要提到(图3):Tg(mpx:eGFP)和Tg(lyz:dsRed),它们分别表达由中性粒细胞特异性启动子髓过氧化物酶和溶菌酶驱动的荧光eGFP或dsRed;Tg(mpeg1:eGFP),由mpeg1启动子激活的标记巨噬细胞;Tg(lcr:eGFP),由球蛋白调控序列驱动eGFP的表达,使红细胞发出绿色荧光。
此外,另外两个系也可用于研究红细胞生成:Tg(gata1:DsRed),这可以使红细胞可视化,因为它们表达由gata1启动子驱动的DsRed;Tg(runx1:GAL4)及其标记的HSPCs。
基因敲入系Tg(asc:asc- egfp)允许炎症小体组装的实时单细胞可视化,而基因敲入系Tg(uas:ascΔCARD-eGFP)和Tg(uas:gpb4)可以分别抑制或激活特定细胞类型的炎症小体。

图3. 斑马鱼转基因系在炎症和炎性小体在造血中的研究。使用这些细胞系可以实现特定的造血细胞类型和炎症小体组装的实时可视化。化学和遗传筛选方法也适用于斑马鱼,在高通量筛选和体内成像方面具有无脊椎动物模型的某些优势,如高繁殖力、光学透明度、快速和外部发展等。缩略语:ASC,凋亡相关斑点样蛋白,含CARD;造血干细胞,造血干细胞和祖细胞;PYD,吡喃结构域;UAS,上游激活序列。
特异性启动子分别为髓过氧化物酶和溶菌酶;Tg(mpeg1:eGFP),由mpeg1启动子激活的标记巨噬细胞;Tg(lcr:eGFP),由球蛋白调控序列驱动eGFP的表达,使红细胞发出绿色荧光。
此外,另外两个系也可用于研究红细胞生成:Tg(gata1:DsRed),这可以使红细胞可视化,因为它们表达由gata1启动子驱动的DsRed;Tg(runx1:GAL4)及其标记的HSPCs。
突变系Tg(asc:asc- egfp)允许炎性小体组装的实时单细胞可视化,而突变系Tg(uas:ascΔCARD-eGFP)和Tg(uas:gpb4)可以分别抑制或激活特定细胞类型的炎性小体。
六、斑马鱼模型阐明了炎性小体在造血中的重要性
如前所述,炎症小体激活和造血之间的关系已被提出用于几种医疗条件;例如,造血系统偏向与慢性炎症疾病有关。
然而,其机制和信号通路尚不清楚。这方面的第一批证据之一是证明了人的GATA1在体外可以被许多caspases切割,而在人的红细胞前体中可以被caspase-3在体内切割。
此外,在小鼠HSPCs中Nlrp1a的激活引发了这些细胞的热解死亡和白细胞减少,而在化疗或病毒感染诱导的造血应激期间,Nlrp1a的激活导致了长时间的细胞减少、骨髓发育不全和免疫抑制。此外,在嘌呤能信号、补体激活和G-CSF给药过程中,theNLRP3炎症小体对于从BM中释放小鼠热休克蛋白是必需的。
值得注意的是,小鼠移植实验表明,NLRP3敲除,无论是在HSPCs中还是在BM基质细胞中,都会导致HSPC的归位和移植受损。
此外,已发现NLRP3炎性小体在骨髓增生异常综合征患者中驱动克隆扩增和细胞焦亡。因此,已经证明,虽然典型炎性小体对于斑马鱼幼鱼HSPC的出现是必不可少的,但它在两个不同的水平的稳态条件下自主调节HSPC的分化:红髓细胞命运决定和终末红髓分化(图1和表1)。
因此,不同炎性小体的遗传抑制作用,例如Gbp4[Tg(uas:gpb4)]或ASC Tg(uas:ascΔCARD eGFP)和caspase a(Caspa)的药理学抑制(斑马鱼的功能相当于哺乳动物的CASP1)导致中性粒细胞和巨噬细胞数量减少,同时伴随着幼鱼红细胞数目的增加反之,通过mRNA注射强制同时表达ASC和Caspa,增加了中性粒细胞数量。
用甲硝唑前药对中性粒细胞表达细菌硝基还原酶的幼鱼进行中性粒细胞消融实验,进一步证实了上述结果;其中caspase-1 Caspa的药理抑制完全损害了消蚀后中性粒细胞的恢复。进一步分析显示Tg(runx1:GAL4)中HSPC的出现未受影响,提示炎性小体可调控HSPC分化。
机制上,炎性小体激活HSPCs导致Gata1蛋白数量减少,促进骨髓生成,抑制红细胞生成。这一机制似乎在进化上是保守的,因为来自SPI1 eYFP和GATA1 mCherry的报告小鼠的HSCs中CASP1的药理学抑制迅速增加了GATA1蛋白量,而不影响SPI1的蛋白量,导致以髓系分化为代价的红系分化增加(图1)。
此外,人红白血病系K562的红系分化也因抑制CASP1而受损,但CASP4不受影响,这表明典型炎性小体通过抑制GATA1蛋白的数量在红系末端分化中发挥作用。体外实验证实了重组人CASP1能够在D300残基处切割GATA1。
综上所述,这些结果支持了这样一种假设,即在HSPCs中,GATA1蛋白的数量由典型炎性小体微调,以调节红髓决定,同时在红髓祖细胞中,调节其终末分化。
此外,通过HSPC标记物runx1/cmyb和Tg (cd41:eGFP)幼鱼的整体原位杂交检测,炎性小体和巨噬细胞来源的Il1b似乎是斑马鱼胚胎HSPC扩增所必需的。然而,需要进一步的研究来阐明在稳态中激活HSPCs中的炎症小体的信号,以及所涉及的炎性小体成分(见未解决的问题)。
GATA1和CASP1之间的这种密切关系在临床水平上是相关的,因为造血系的偏置与具有炎症成分的疾病的较高发病率相关,如感染、类风湿关节炎、动脉粥样硬化、炎症性肠病、肾脏和肺部疾病。自身免疫性疾病,神经退行性疾病和某些癌症这类疾病的一个例子是中性粒细胞性皮肤病,以中性粒细胞在患者多形性皮损中积累为特征。
一种缺乏丝氨酸肽酶抑制剂的斑马鱼系,Kunitz 1a型(Spint1a),已被用作该疾病的模型,因为它显示中性粒细胞增多、皮肤损伤和高皮肤中性粒细胞浸润。因此,在这个模型中,使用CASP1/Caspa抑制剂Ac-YVAD-CMK抑制典型炎症小体,可以恢复Gata1的数量并减少中性粒细胞的数量,尽管皮损和中性粒细胞浸润都没有影响(图4)。
另一种典型炎症小体抑制可能被认为是有待进一步探索的治疗靶点的疾病是Diamond-Blackfan贫血(DBA),这是一种遗传性疾病,由于GATA1蛋白数量减少导致红细胞生成受损。我们通过使用特定的morpholino减少Gata1的量来模拟斑马鱼的DBA。
在DBA模型中,Ac-YVAD-CHO对Caspa的抑制可以部分挽救贫血,并增加Gata1和降低Spi1蛋白的数量(图4)。
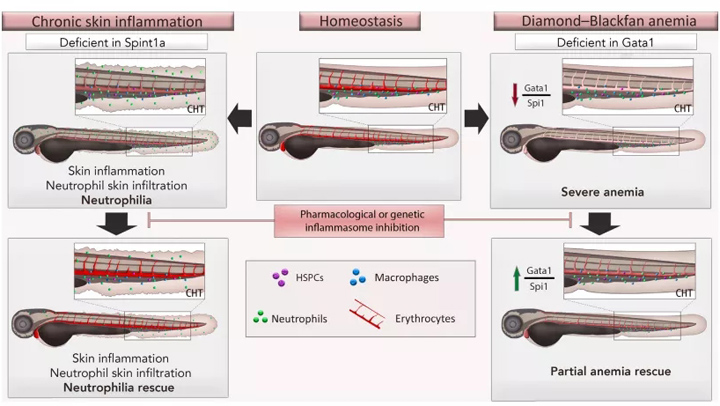
图4. 用于研究造血的斑马鱼疾病模型。在稳态条件下,胚胎造血发生在受精后2-3天的尾侧造血组织(CHT)(中图)。Spint1a缺陷系(spint1ahi2217Tg/hi2217Tg)是研究与慢性炎症相关的造血系统偏向的一个极好的模型,因为它很容易观察到中性粒细胞(左图)。Gata1a变种动物是一种可适应的菱形-黑扇贫血模型。在这两种模型中,抑制炎性小体可以挽救造血系统偏向。简称:造血干细胞,造血干细胞和祖细胞。
总之,在这些斑马鱼疾病模型中,药理抑制炎症小体可以挽救中性粒细胞炎症和贫血;这表明炎症小体可以在某些慢性炎症疾病和DBA的造血谱系偏差中发挥主要作用,至少是源于斑马鱼模型。尽管如此,这些发现确实值得进一步关注,在某些情况下,为潜在的治疗干预确定新的药物靶点。
七、结论
斑马鱼模型在生物医学研究中具有独特的优势,在这里,我们讨论了它与设计研究的相关性,以更好地理解炎性小体激活的作用和机制。慢性炎症疾病模型的可用性,包括中性粒细胞性皮肤病和罕见疾病,如DBA,提供了一个优秀和独特的工具,以快速确定一些这些病理的新治疗方法,而且,用于筛选临床批准的药物库,可能有助于相对快速地重新利用。
此外,功能强大的CRISPR-Cas9技术可以快速产生候选致病突变,甚至可以在F0代进行分析,而无需产生耗时的缺陷系,以确认其致病性,并进行靶向或先导药物筛选。
必须找到参与造血的特异性炎性小体,为慢性炎症疾病相关的造血系统偏向开发更特异性的治疗方法(见突出问题)。在体内追踪造血细胞的可能性,以及可用的炎性小体,包括实时可视化其组装的报告,可以使该模型与其他脊椎动物模型互补,从而研究造血和炎症之间的关系,特别是炎性小体与造血之间的关系。
参考资料:
[1] Palis, J. and Yoder, M.C. (2001) Yolk-sac hematopoiesis: thefifirst blood cells of mouse and man. Exp. Hematol. 29, 927–936
[2] McGrath, K.E. et al. (2011) A transient defifinitive erythroid lineagewith unique regulation of the β-globin locus in the mammalianembryo. Blood 117, 4600–4608
[3] Bertrand, J.Y. et al. (2007) Defifinitive hematopoiesis initiatesthrough a committed erythromyeloid progenitor in the zebrafifishembryo. Development 134, 4147–4156
[4] Jagannathan-Bogdan, M. and Zon, L.I. (2013) Hematopoiesis.Development 140, 2463–2467
文丨Lola Rodríguez-Ruiz,Juan M. Lozano-Gil,Christophe Lachaud,Pablo Mesa-del-Castillo,María L.
Cayuela,3 Diana García-Moreno,Ana B. Pérez-Oliva and Victoriano Mulero
翻译丨郭盟
DOI:Trends in Immunology, Month 2020